诺奖得主 Jennifer Doudna 团队,又在顶刊《科学》上投下了一枚重磅冲击波。
这一次,他们展示了一项“超越自然演化”的成果:基于AI设计的蛋白质,创造出的新型基因编辑蛋白SynTnpB,部分变体的活性,甚至直接对标——不,是超越了天然酶。

论文链接:http://www.science.org/doi/10.1126/science.aed6123
冷冻电镜的结果进一步证实了这一点:这些工程化蛋白,在不同的构象状态下,于RNA-DNA界面形成了全新的、稳定的相互作用。这不仅是技术上的突破,更是首次通过实验,实锤了AI设计的RNA引导核酸酶的结构。
研究团队指出,当生成式生物学与从头设计方法联手,未来突破自然序列的边界,扩展这类蛋白的功能和设计空间,就不再是天方夜谭。
超越自然的AI蛋白质设计
众所周知,CRISPR-Cas技术已是基因编辑领域的明星,能精准切割DNA或RNA。但要凭空设计一个自然界中不存在、却依然保持活性的RNA引导核酸酶,谈何容易。现有的序列模型,生成出来的酶往往还是太“像”天然的了;而结构引导设计,虽然能探索更大的序列空间,却依然难以在大幅改写序列的同时,保住那份关键的活性。
这一次,Doudna团队没有走寻常路。他们提出了一种全新的策略:将结构与进化信息结合起来,指导AI进行蛋白设计。目标是生成那些序列高度分化、但核心活性依然在线的RNA引导核酸酶。
为了验证这个策略,他们挑选了一个“小个子”——小型RNA引导核酸酶TnpB作为实验对象。整个过程,清清爽爽,分两步走:
1.设计模块:生成全新的TnpB序列
他们首先祭出ESM逆折叠模型(ESM-IF1),让它根据TnpB的三维结构,从头“写”出候选序列。这些AI生成的序列,能保留TnpB的整体折叠形态和RuvC结构域的关键催化位点(DED催化三联体);但问题也随之而来——它们也会擅自改动部分负责识别RNA/DNA的残基。
为了解决这个“bug”,团队基于进化信息构建了一个“序列掩码”:那些位置保守性(Ci)极高,或者与核酸序列存在共同进化耦合强度(σi)极高的残基,被强制锁定为野生型,不允许改动;剩下的位置,则交给ESM-IF1自由发挥,重新设计。
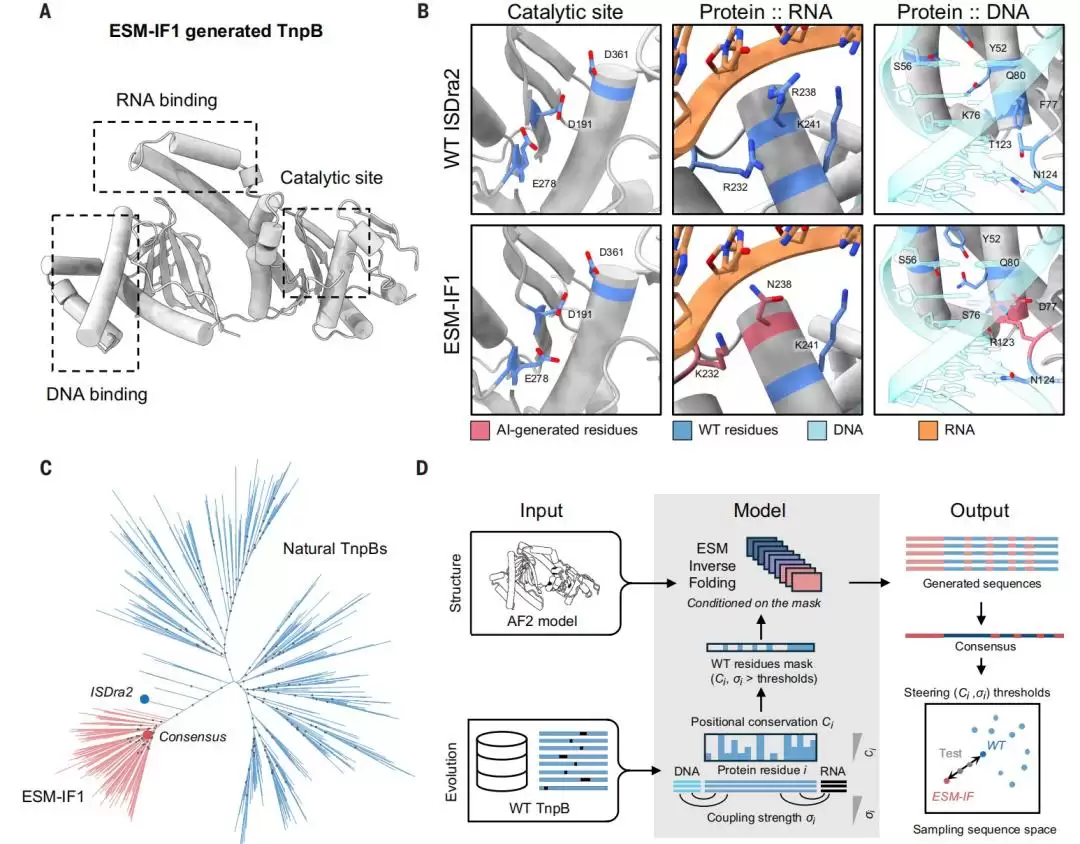
图|使用ESM逆折叠(ESM-IF1)模型进行RNA引导核酸酶TnpB设计的策略概览。
2.筛选模块:从海量变体中“淘金”
序列设计完成,只是第一步。接下来,团队用了一个巧妙的细菌ccdB毒素系统来筛选具有活性的变体:只有那些能切割毒素质粒的TnpB变体,才能让细菌活下来。
最初,仅用Ci约束进行全长设计,活性表现一般。于是他们决定拆开来看,分别测试TnpB的两个结构域:REC(主要参与DNA识别)和NUC(与RNA结合和催化相关)。结果发现,NUC结构域更能耐受序列变化。基于这个发现,他们采用Ci/σi双条件约束组合,在1980个REC-NUC组合中,最终筛出了9个高活性、高多样性的“尖子生”。

图|基于位置保守性(Ci)和耦合强度(σi)阈值条件化生成的AI变体SynTnpB的筛选流程。
活性超越天然酶
整体来看,AI设计的这些SynTnpB变体,不仅在细菌里表现良好,在植物和人类细胞中也保留了活性,甚至部分变体的表现超越了天然酶。冷冻电镜结构进一步揭示了背后的秘密:这些AI设计出的变体,能在不同构象状态下,像“胶水”一样稳定住RNA-DNA界面。具体来看:
1.在人类和植物细胞中“大显身手”
筛选出的变体,在人类和植物细胞中均保留了编辑活性,部分变体甚至超过了天然酶。 团队将9个变体分别放入HEK293T细胞(人类)和拟南芥原生质体(植物)中进行验证。在人类细胞中,多数变体的活性与野生型相当,其中v1和v5表现尤其亮眼,最高编辑率达到50%,大约是野生型的1.8倍。而在植物细胞实验中,v1同样在多数被测试的靶点上表现出优于野生型的性能。
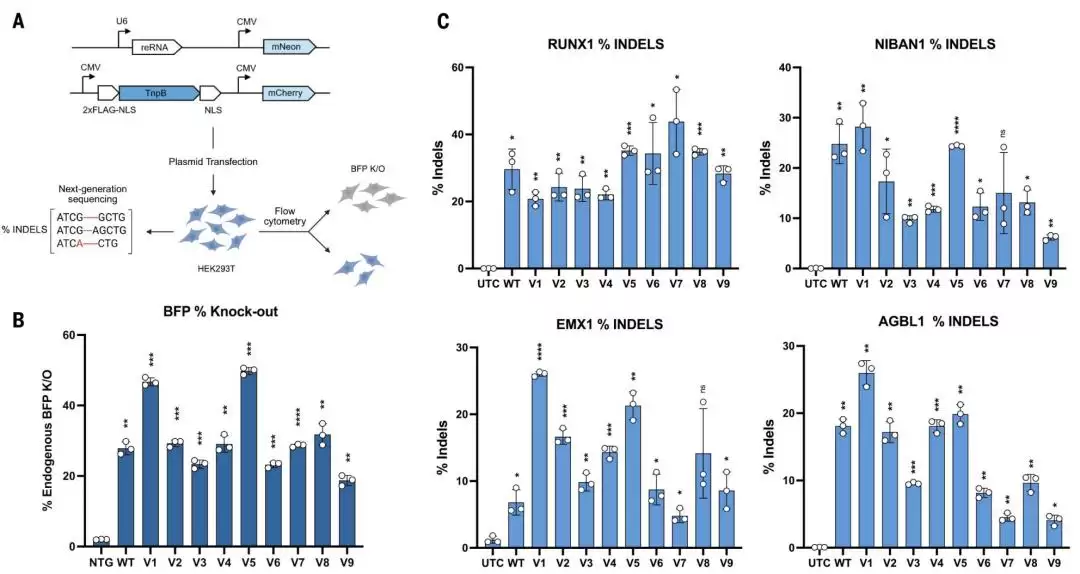
图|AI生成的TnpB变体在HEK293T细胞中的基因组编辑效果。
2.特异性和“脾气”摸了个底
从特异性来看,SynTnpB变体保留了典型的TTGAT TAM偏好,但脱靶表现则“变体各异”。全基因组Tn5标签化分析显示,v1的特异性与野生型持平,而v5和v7则检测到了更多的脱靶位点。
至于表达和生化特性,编辑活性的差异并不能简单归因于表达水平。Western blot结果显示,各变体的表达水平与编辑活性之间没有直接关联。以v7变体为例,它的热稳定性与野生型相当,也保留了较强的靶链切割能力,但切割反应的整体速度比野生型要慢一些。
3.冷冻电镜下的“实锤”
冷冻电镜的结果令人振奋。AI生成的残基,在多个结构域与RNA-DNA链形成了新的接触点,稳定了不同构象状态下的界面。更令人惊喜的是,他们还捕获到了一种此前在TnpB中从未被报道过的TAM结合构象的中间状态。
团队选择了序列分化程度较高的v7变体进行冷冻电镜解析。这个变体与野生型的序列一致性为77%,在RUNX1位点显示出较高的编辑效率。实验结果显示,冷冻电镜结构分辨率达到了2.8 Å,并成功捕获了两种构象:TAM结合状态和R-loop形成状态。
在TAM结合状态中,蛋白和RNA的构象与ISDra2 TnpB二元复合物相似,但此时已结合了DNA,并在异源双链的种子区形成了单碱基配对。AI生成的N4R残基与保守的“磷酸锁”残基K84共同稳定了这一配对。这种构象中间体,此前从未在TnpB中被观察到。
而在R-loop形成状态中,reRNA和靶DNA已经形成了比较完整的配对结构。此时,AI生成的残基分布在REC、NUC和lid等多个区域,与这段RNA-DNA双链紧密接触;同时,桥接螺旋仍保留了有助于双链形成的弯折运动。
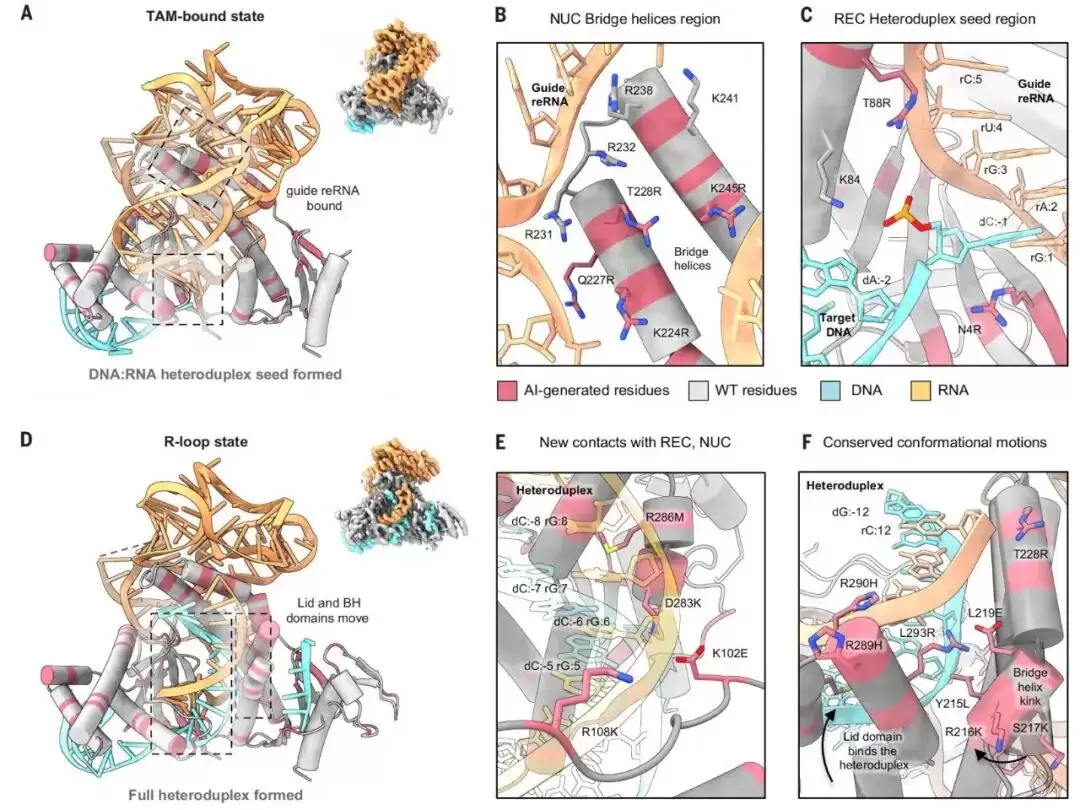
图|AI生成变体v7的冷冻电镜结构,揭示了RNA-DNA界面残基和保守的构象运动。
不足与展望
当然,这项研究并非完美无缺。研究团队也坦诚地指出了几个关键问题。
首先,在设计目标上,TnpB的不同结构域对序列改变的耐受性存在差异。与DNA识别相关的REC结构域就相当“敏感”,对替换反应较大;而负责RNA结合和催化的NUC结构域则“包容”得多。未来,这种模块化设计原则是否适用于其他多结构域蛋白,还需要进一步验证。
其次,在数据依赖上,序列掩码的方法高度依赖于足够丰富的进化信息,对数据库中序列的多样性和代表性要求很高。更丰富、更均衡的蛋白-核酸配对数据,是推广这一方法到其他多状态蛋白或核酸结合蛋白的关键。
最后,在机制理解上,AI生成的残基究竟是如何影响核酸结合和构象变化的,这背后还有大量谜题等待解开。研究团队表示,这项研究中冷冻电镜捕获到的、此前未被报道的TAM结合状态,可能代表了一种动力学中间体,这为未来研究RNA引导核酸酶的瞬时构象,打开了一扇新的大门。
更多技术细节,详见原论文。